Эволюция вприпрыжку
Эволюция вприпрыжку
Михаил Бурцев
Один из краеугольных камней современной теории эволюции — идея о ее градуальности. В соответствии с ней, отбор последовательно закрепляет мутации, каждая из которых слабо влияет на фенотип. Шаг за шагом этот процесс приводит к появлению сложных адаптаций и разнообразию видов. Теория говорит, что для того, чтобы такая пошаговая эволюция имела место, необходима адаптивность каждой мутации или хотя бы ее нейтральность. Однако, теория Мото Кимуры, разработанная в 1985 г., предлагает дополнительный альтернативный сценарий.
Кимура рассмотрел случай, когда приспособленность организма зависит от нелинейного взаимодействия разных генов. При взаимодействии разных вариантов (аллелей) двух генов может возникнуть ситуация когда невозможно эволюционировать пошагово из одного состояния в другое. Допустим, что в одном гене возможны два варианта (A и a) а в другом гене вариант (B и b). Если приспособленность индивидуумов с генотипом AB и ab высокая, а состояние генотипов Ab и aB приводят к низкой приспособленности. Тогда, получается, что для эволюции от варианта АB к аb необходимо совпадение мутаций сразу по обеим копиям гена, так как если мутация будет одна, то адаптивность снизится. Однако, Кимура заметил, что если рассматривать не отдельную особь, а популяцию в целом, то вероятность прыжка через долину с низкой приспособленностью увеличивается. Причина в том, что ситуация, когда популяция состоит только из особей с генотипом АB маловероятна, обычно в ней присутствует некоторое число мутантов Аb и аB. А в этом случае переход от АB к аb может произойти без фиксации промежуточного фенотипа с худшей приспособленностью.
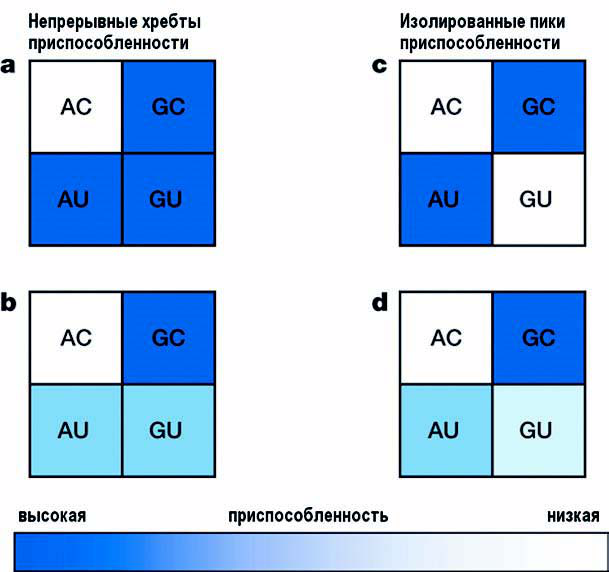
Рис. 1. Переключение между Уотсон-Криковскими парами AU и GC может происходить на плоском (а) или восходящем (b) ландшафте с непрерывными хребтами приспособленности или через долину, образуемую промежуточными вариантами с одинаково © или по-разному (d) пониженной приспособленностью (рисунок из обсуждаемой статьи в Nature).
В работе [1] исследователей из Центра геномной регуляции в Барселоне, Испания и Университета Мичигана, США изучалась эволюция митохондриальной транспортной РНК. Они рассмотрели переключения между Уотсон-Криковскими парами AU и GC (см. рис. 1) и показали, что промежуточные варианты GU и AC связаны с уменьшением приспособленности. Полученные для межвидовой и внутривидовой вариации результаты находятся в соответствии с гипотезой Кимуры о компенсаторной эволюции. Подобные компенсаторные переключения, из AU в GC, или наоборот, составляют основную долю от всех замен во вторичный структурах мт-тРНК, что показывает, что эти молекулы эволюционирует не шаг за шагом, как предсказывает классическая теория, а почти одновременной фиксацией двух отдельных мутаций. Исследователи говорят, что «Несмотря на наглядность пошаговой модели адаптивной эволюции, кажется, что для мт-тРНК большинство замен зависят от популяционной динамики, позволяющей перепрыгивать к лучшей приспособленности».
«Наша работа показывает, что невозможно описать эволюционный процесс целого класса молекул (митохондриальных тРНК) без понимания, как различные полиморфизмы взаимодействуют друг с другом в одной популяции», — говорит Фёдор Кондрашов, основной автор статьи и руководитель группы Эволюционной геномики в Центре геномной регуляции. «Наши результаты показывают, что некоторые мутации, уменьшающие приспособленность, не обязательно являются эволюционными тупиками. Наоборот, две по отдельности вредные мутации, которые в комбинации дают полезный эффект, открывают новые пути для дальнейшего движения эволюции», объясняет Кондрашов.

Федор Кондрашев
Федор Кондрашов ответил на вопросы ТрВ.
— Как возникла идея провести это исследование?
— Из нашей предыдущей работы [2] следовало, что компенсаторная эволюция, то есть переходы из одного Ватсон-Криковского состояния в другое происходят очень часто. Но мы тогда не смогли разобраться, как именно такие переходы происходят.
— Насколько часто в эволюции может встречаться подобный тип переходов?
— Конкретно для вторичных структур мт-тРНК до 60% от всех замен, то есть очень значительный кусок. Насколько этот эволюционный механизм распространён в других молекулах абсолютно непонятно. Основная новизна нашей работы заключается в том, что нам впервые удалось показать, что хоть в каких-то случаях такой не пошаговый механизм является важным.
— Можно ли обобщить результат на комбинации аллелей разных эпистатически взаимодействующих генов?
— Пока что нет, и, наверное, это является самым интересным вопросом на следующие 10–20 лет. Пока совершенно непонятно насколько эпистатические взаимодействия, возникающие между разными аллелями в одной популяции, важны на макроэволюционном уровне. Единственное исключение — мт-тРНК.
— Можно ли как-то это проверить?
— Активно думаем на эту тему. С мт-тРНК мы смогли проверить из-за мутационного неравновесия двух нитей ДНК в митохондрии. Как это проверить в белках или в ядерном геноме пока не понятно.
— Планируется ли развивать эту работу? Если да, то как, если нет, то что на очереди?
— Было бы забавно посмотреть, насколько важен такой процесс в эволюции белков. К сожалению, в белках гораздо сложнее рассмотреть взаимодействия всего двух сайтов (локусов) так как аминокислотные остатки часто взаимодействуют не попарно. Так же актуально дальнейшее развитие теории Кимуры — было бы забавно посмотреть насколько эпистатические взаимодействия между многими генами может повлиять на эволюцию через долины приспособленности.
Михаил Бурцев
[1] Margarita V. Meer, Alexey S. Kondrashov, Yael Artzy-Randrup & Fyodor Kondrashov (2010) «Compensatory evolution in mitochondrial tRNAs navigates valleys of low fitness». Nature. DOI: 10.1038/nature08691.
[2] Kern AD, Kondrashov FA. Mechanisms and convergence of compensatory evolution in mammalian mitochondrial tRNAs. Nat Genet. 2004 Nov;36(11):1207–12.
Более 800 000 книг и аудиокниг! 📚
Получи 2 месяца Литрес Подписки в подарок и наслаждайся неограниченным чтением
ПОЛУЧИТЬ ПОДАРОКЧитайте также
Хромосомы и эволюция
Хромосомы и эволюция Точечные мутации ДНК не могут служить причиной возникновения новых видов животных, так как виды отличаются один от другого не столько последовательностью нуклеотидов в ДНК, сколько количеством и размером хромосом (в которых находится ДНК). Новые
Эволюция продолжается?
Эволюция продолжается? Когда говорят об эволюции, в сознании большинства сразу всплывают страницы учебника биологии, на которых изображен ряд постепенно распрямляющихся фигур, наглядно демонстрирующий «происхождение вида путем естественного отбора». Видимо, из всего
Эволюция коммунизма
Эволюция коммунизма Советский коммунизм сложился не сразу. И он изменялся со временем. Можно констатировать такие периоды в его истории: зарождения, юности, зрелости, кризиса и краха. Первый период охватывает годы от Октябрьской революции 1917 года до избрания Сталина
Эволюция капитализма
Эволюция капитализма Капитализм не есть нечто раз и навсегда данное. В его истории различают два периода – периоды «старого» и «нового» капитализма. Я их различие вижу в следующем.«Старый» капитализм был по преимуществу множеством индивидуальных капиталов, вкрапленных
Иная эволюция[314]
Иная эволюция[314] Понятие эволюции может охватывать радикально отличающиеся друг от друга явления. Если, например, в книге «Сумма технологии» я когда-то писал о двух разных эволюциях, то я имел в виду эволюции биологическую и технологическую. Для биологической характерна
Иная эволюция[314]
Иная эволюция[314] Понятие эволюции может охватывать радикально отличающиеся друг от друга явления. Если, например, в книге «Сумма технологии» я когда-то писал о двух разных эволюциях, то я имел в виду эволюции биологическую и технологическую. Для биологической характерна
Эволюция бандита
Эволюция бандита Советские гангстеры появились в годы новой экономической политики (1921—1929), когда большевики вынуждены были в целях выживания системы, серьезно обескровленной в период Гражданской войны (1918—1922), начать совмещать плановую экономику с рыночной. В
Эволюция forever
Эволюция forever Опрос британских школьников выявил удивительные факты.Выяснилось, что телефон изобрела ныне действующая королева. По крайней мере так думает каждый десятый. Оказалось также, что Ньютон открыл огонь. Не в том смысле, в каком открывает его артиллерия по врагу,
Государство и эволюция
Государство и эволюция Мы имеем сколько-то связные представления о человеке в Истории только до тех глубин времен, когда была уже письменность и изрядная архитектура, но не раньше. А ими сопровождалось существование государства.Человеком догосударственным занимаются
Эволюция государства
Эволюция государства Примем, что «целью» государства является выполнение функции, обеспечивающей жизнеспособность государства (назовем ее целью № 1). Это означает, что функция должна выполняться в некотором (довольно широком) диапазоне условий. Информация, необходимая
Эволюция номер два
Эволюция номер два Верующие люди убеждены, что всё живое на Земле создал Бог, и только ему принадлежит право на то, чтобы «вдыхать» жизнь в мертвую материю. Однако с возникновением цивилизации человек неоднократно бросал вызов Создателю, претендуя на то, чтобы сравниться
5. Эволюция ритуала
5. Эволюция ритуала Итак, возможность кодирования всей необходимой информации о любой структурированной деятельности в формах некоего заместительного движения сомнений не вызывает. Высказанные здесь соображения касаются только механизма ее передачи какому-то другому
Эволюция лошади
Эволюция лошади Почти в каждом учебнике, там, где речь идет об эволюции, присутствует схема эволюции лошади. Схема показывает, что современная лошадь произошла от четвероногого существа размером с кролика. Этот процесс, занял миллионы лет (рис. 62). Схема была составлена на
Эволюция и наука
Эволюция и наука Эволюционная теория не является строго научной, она может быть квалифицирована лишь как гипотеза.Чтобы теория, вырастая из рабочей гипотезы, могла считаться научной, должно быть экспериментальное подтверждение ее постулатов. Другими словами, мы должны
Эволюция человека
Эволюция человека Точно такое же заключение можно сделать и об эволюции человека. Несмотря на догматические утверждения эволюционистов о том, что человек произошел от обезьяноподобных существ, если тщательно и объективно изучать ископаемые доказательства, будет
Эволюция познания
Эволюция познания Библиоман. Книжная дюжина Эволюция познания Уильям Арнтц, Бетси Чейс, Марк Висенте. Кроличья нора, или Что мы знаем о себе и Вселенной / Пер. с англ. А. Степановой. – М.: Эксмо, 2011. – 448?с. – 10 100?экз. Зачем сытой и аккуратно одетой части человечества (а